 m.quanpro.cn
m.quanpro.cn
媒体:原创 作者:国家植物园
专业号:国家植物园
2023/3/29 17:41:35
分网
土壤碳封存——我们脚下的碳中和方案 | 芳兰葳蕤
土壤中的碳循环,被《科学》杂志誉为“最后的前沿”。缓解气候变化的可持续解决方案,就在植物与微生物相互博弈的湿地、雨林、草原…就在我们脚下每一寸土壤的碳库中。
冯晓娟
国家植物园、中国科学院植物研究所副所长 研究员
众所周知,在气候变化的背景下减少温室气体的排放,实现碳中和是全球许多国家向联合国作出的承诺。提起碳中和,大家可能会想到节能减排,但是大家知道吗?每年自然生态系统中植物和土壤的系统和大气二氧化碳的交换通量是人为排放量的十倍以上。特别是我们脚下的土壤,是植被碳库的3~4倍。

那么,如何实现土壤碳封存呢?这是一个听上去很简单,但是实际上非常复杂的科学问题。土壤被《科学》杂志誉为最后的前沿。我是冯晓娟,来自中国科学院植物研究所。

在传统的观念中,我们认为植物的凋落物,也就是枯枝落叶是土壤碳的最主要的来源。特别是陆地植物中特有的木质素大分子,当木质素的含量高时,凋落物的降解较慢,也更容易在土壤中积累。所以长期以来大家都认为木质素是导致土壤碳积累的一个主要的贡献者,很多的土壤碳库模型里也将木质素作为一个表征稳定碳库的重要参数。
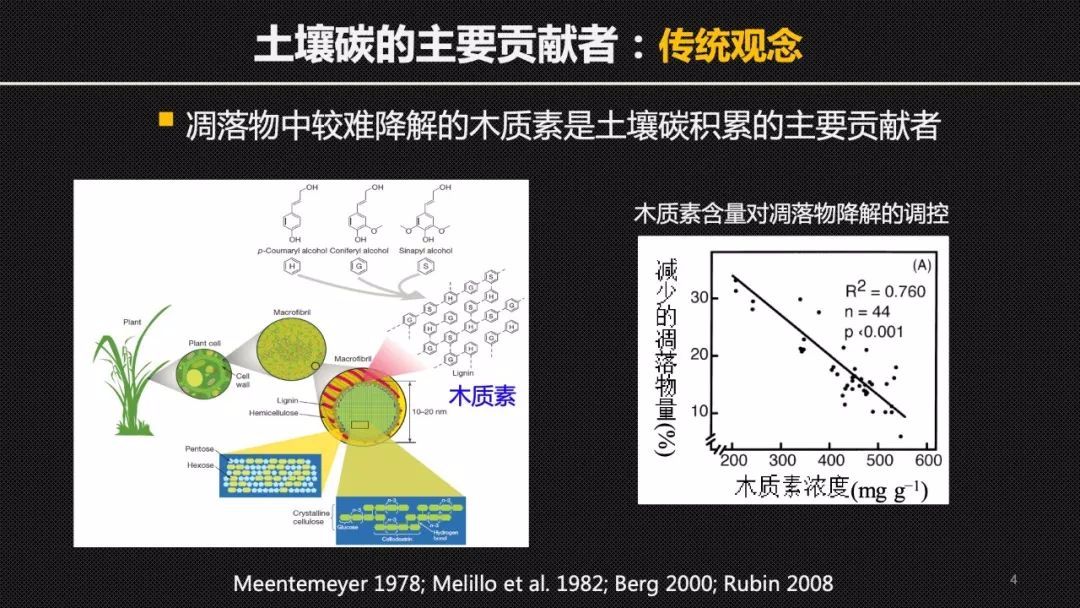
那么这里就出现了一个关键的科学问题!想要准确地定量地研究土壤中木质素的降解速率其实是一个不小的挑战,因为土壤有机质的组成实在是太复杂了,我们没有办法去区分那些结构、来源都不同的生物大分子。但是,我们可以借助化学的方法,将复杂的生物大分子击碎,然后去研究它的小分子单体化合物,也就是生物标志物进行碳14的分析。为比较植物或者是其他的组分来源的这样的分子组分在天然环境里的周转提供最直接的证据。
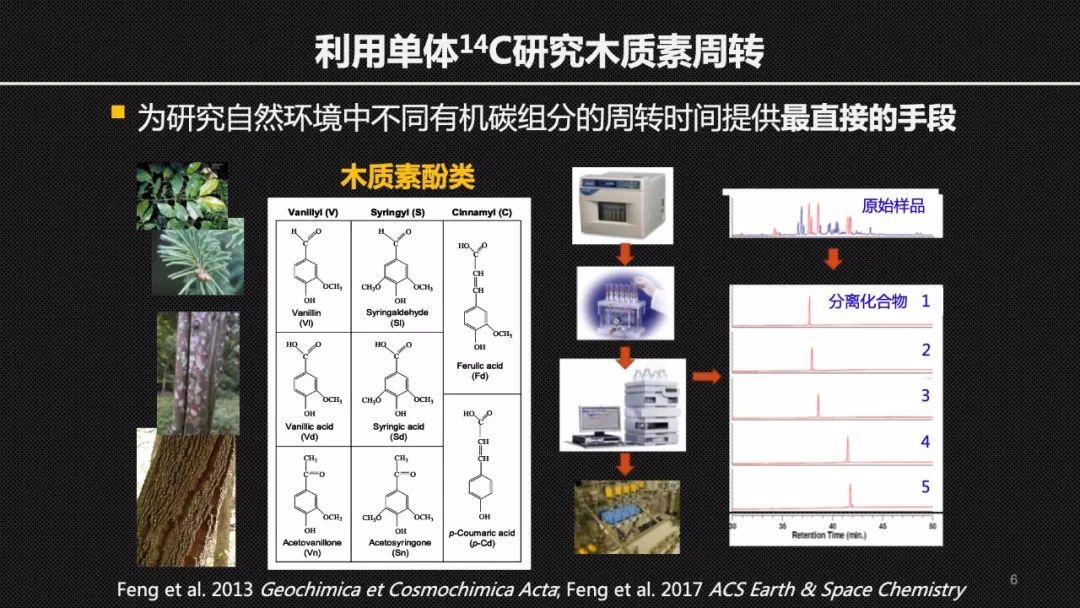
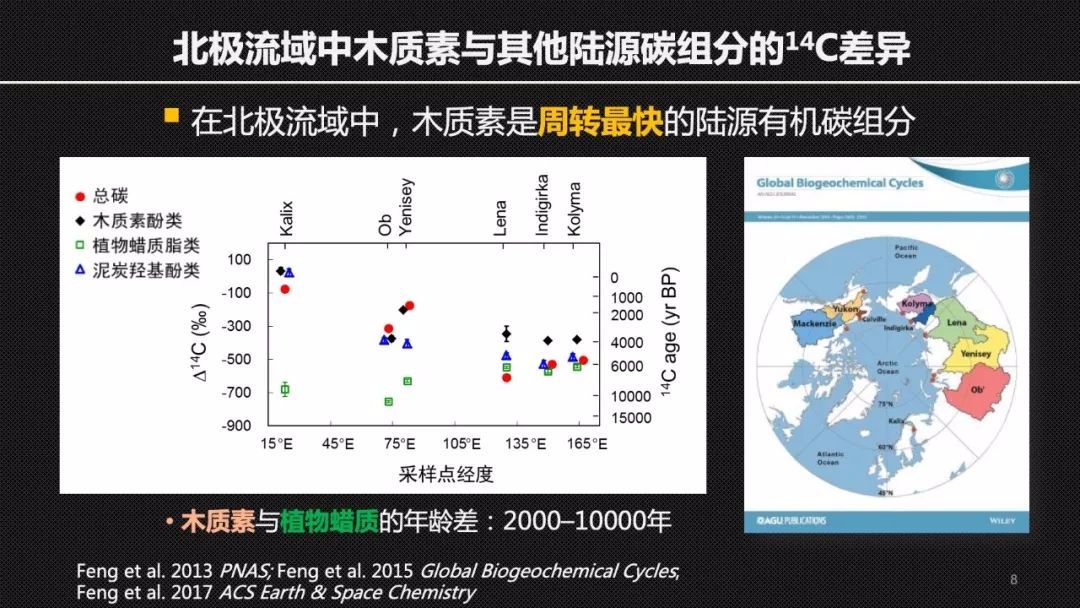
使用单体碳14的方法,我们首先聚焦于北极冻土。这里的土壤有机碳在低温的环境下保存了上万年甚至更长的时间。因此,不同的碳组分可能表现出来截然不同或者差异巨大的周转时间。

我们很惊讶地发现,在北极的几条主要的河流河口沉积物里,木质素是最年轻的来自于陆地植物的有机碳组分。它的碳14年龄和同样来自于陆地植物的植物蜡质脂类相比可以年轻上万年。这就表明在北极的流域里,木质素是周转最快的陆源有机碳组分。这样一个结果颠覆了我们之前的想象,而且也受到了国内外同行的高度评价。
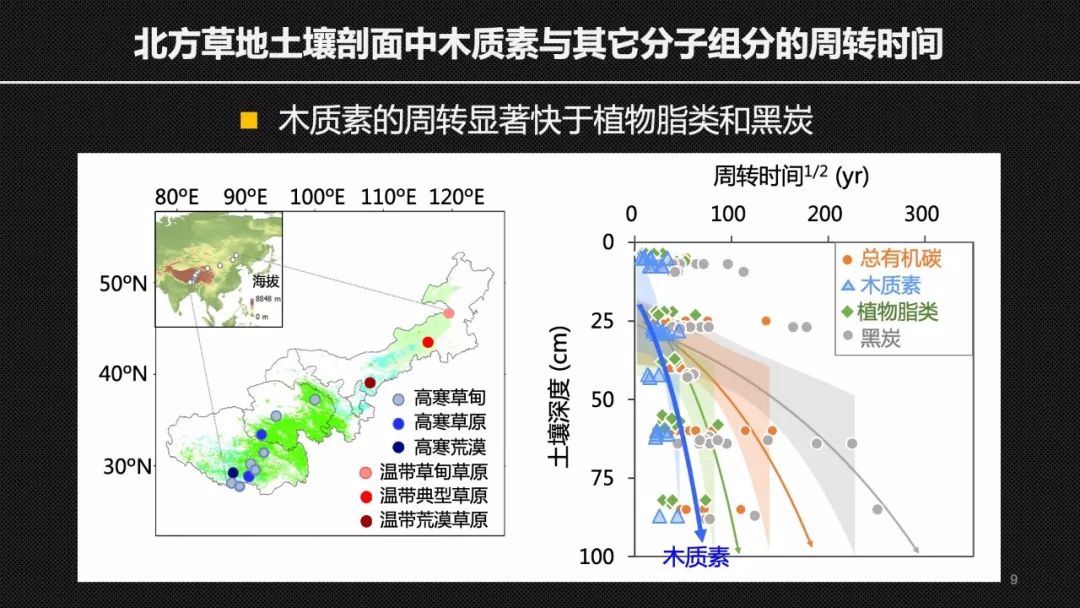
不仅如此,我们进一步将单体碳14的方法应用于我国北方草地跨度超过3000公里的13个土壤剖面中。在这些剖面中我们同样也发现木质素的周转要显著的快于植物的脂类和黑炭。这样一些结果就在大的区域尺度上证明,在矿质土壤剖面以及流域中木质素是快速周转的。为木质素的快速周转提供了关键的科学证据。这也说明木质素对于土壤碳汇的贡献可能被高估。
既然木质素在长时间尺度上没有我们想象中那么稳定,那么究竟是什么组分主导了土壤碳的长期积累?现在最新的观点认为微生物的残体非常重要!在传统的观念中,微生物可能仅仅被看作是有机碳的降解者,而且微生物的活体生物量只占土壤总有机碳库的1%~5%,可以说微不足道。但是,微生物群落的周转非常的快,在它的世代衍替和繁衍中,它的细胞残体以及代谢产物可以在土壤中不断地积累,因此对土壤碳库产生重要的贡献。更重要的是微生物的细胞残体和矿物有紧密的结合,也更容易被矿物保护,所以更加稳定。也就是说微生物作为一个降解者,在降解有机碳的同时,也把一部分容易降解的有机碳转化成为了更难降解的微生物残体,在土壤中积累下来。这样一个由微生物调控的,将活性的有机碳转化和分配到慢性或者惰性碳库中的过程,被形象的比喻成土壤“微生物碳泵”。
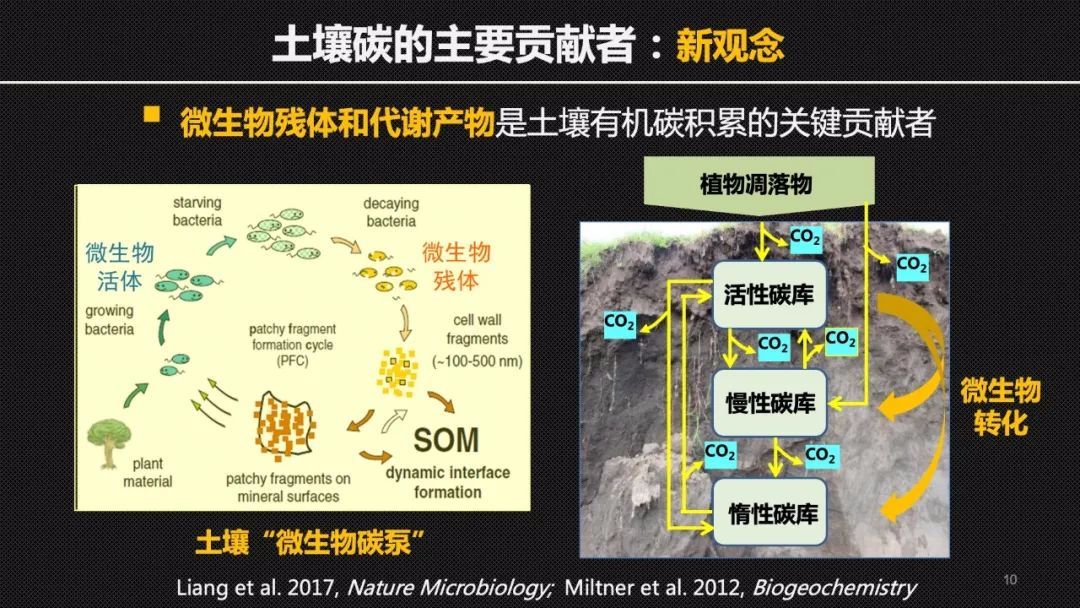
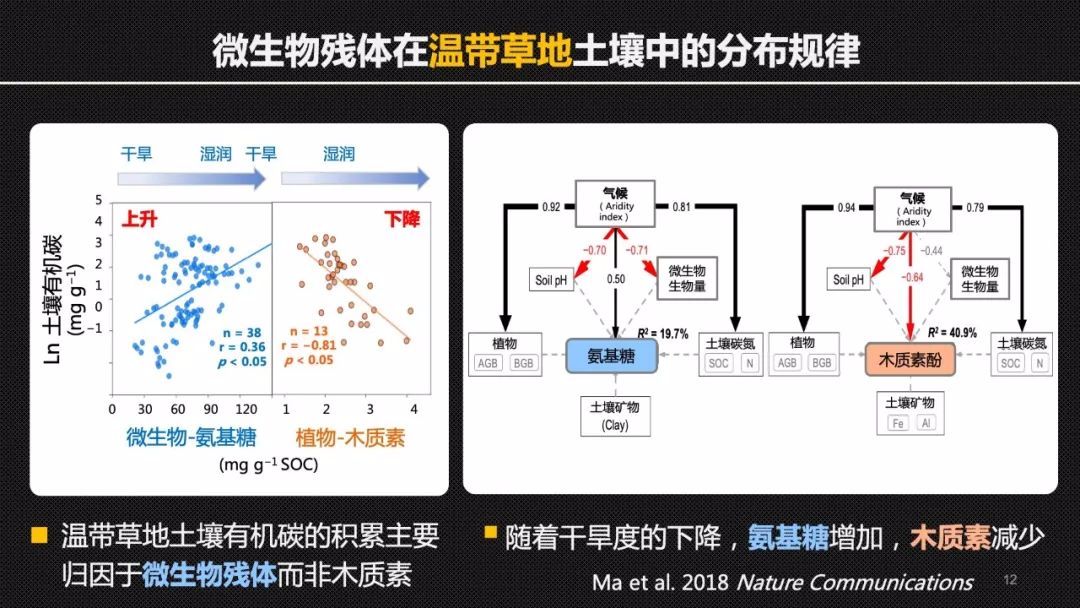
木质素和氨基糖或者说微生物残体此消彼长的这样一个动态变化,还体现在土壤碳库对于增温的响应上。在青藏高原海北站的高寒草地上,我们借助贺金生教授设立的长期增温控水实验平台,对比研究了土壤中不同的分子组分对于增温的响应差异。
贺老师从2011年就开始了这个实验,坚持了十多年。科研人员需要克服高寒恶劣的气候条件,进行严格的气候变化的模拟实验,可以说难度极大。

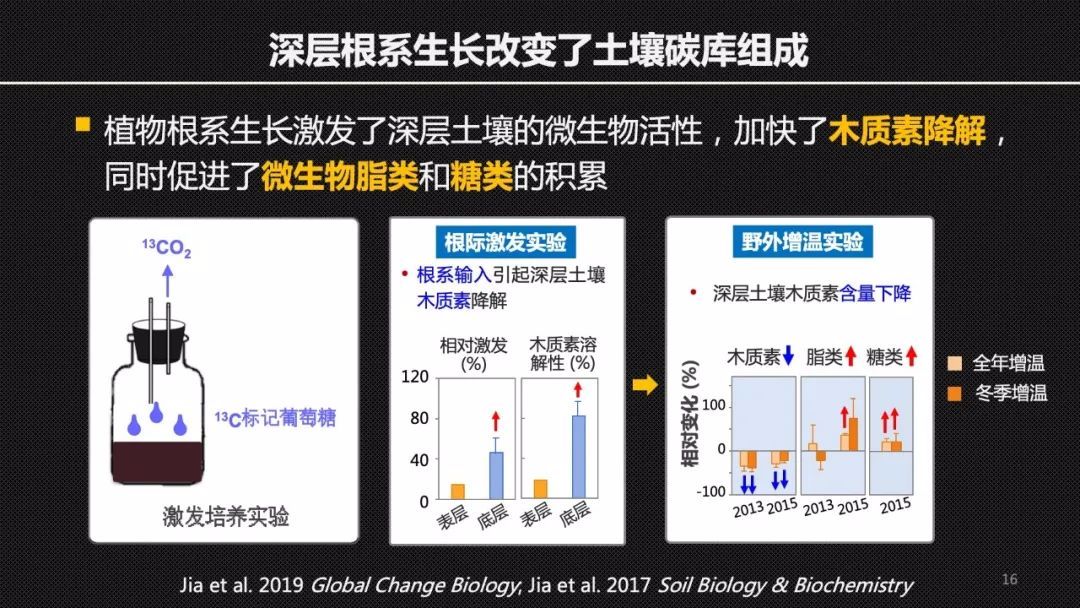
结合海北站32年连续的对于植物群落的这样的一个长期监测的数据,贺老师的研究团队发现,增温和干旱增强了根系比较深的禾草科的植物在整个群落中的竞争优势。

那么这样一个深层根系的生长会如何影响土壤碳的积累或者碳动态呢?我们通过土壤的培养实验,利用碳13标记的葡萄糖来模拟植物根系的碳输入,我们发现新碳的输入激活了深层土壤中微生物的活性,同时加快了木质素的降解。这样一个室内的培养实验验证了野外增温平台上面观察到的结果,也就是在增温的处理下深层土壤中木质素的含量下降。同时我们还发现深层根系的增长,增加了来自微生物的脂类和糖类在深层土壤中的贡献。证明了微生物在土壤碳增汇中的一个关键的作用。它不仅仅是一个降解者,它还可以是土壤碳库的贡献者,这对于我们深入地理解土壤碳循环以及优化碳库模型都有重要的意义。
那么微生物的残体是不是越多越好呢?也并非如此。我们需要意识到土壤“微生物碳泵”是由异养微生物驱动的转化过程,碳泵的运转还需要由初级生产者——植物来提供燃料,也就是植物碳的输入。植物碳输入的量、质以及土壤环境都会对土壤“微生物碳泵”的运转以及效率产生重要的影响。

我们做一个比喻,如果把土壤看作是一个碳的加工厂,土壤碳是我们希望得到的产品,那么植物通过光合作用提供的有机碳的输入就是原材料,而微生物就是加工车间或者加工工人,想要提高产量,我们不仅需要提高对原材料的投入,而且还需要提高加工车间的生产技能或者是生产成效。因此比起微生物残体的总量,我们更应该关心的是土壤“微生物碳泵”的效率,也就是说产生相同的微生物的产品需要消耗多少初级生产力,那些效率比较高的碳泵更有利于土壤碳封存。
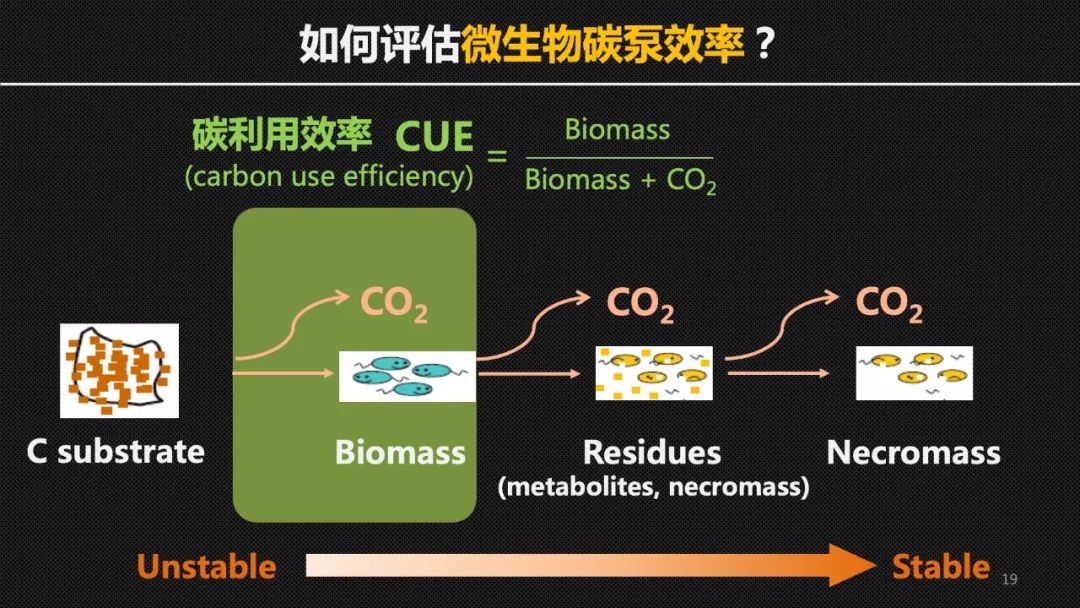
那么如何评估土壤微生物的碳泵效率呢?在微生物的研究中有一个常用的参数叫做微生物的碳利用效率——CUE,它指的是微生物的自身的生物量的增长速率和呼吸速率的比值。通俗地讲就是在相同的食量下,去评估微生物是更容易长肉,还是更容易浪费粮食。但是在微生物死亡以后,仅有一小部的微生物碳或者说残体可以在土壤中积累下来,因此这些微生物残体的保存可能还受到土壤矿物保护等非生物过程或者参数的调控,可能和微生物的CUE生理指标解耦。
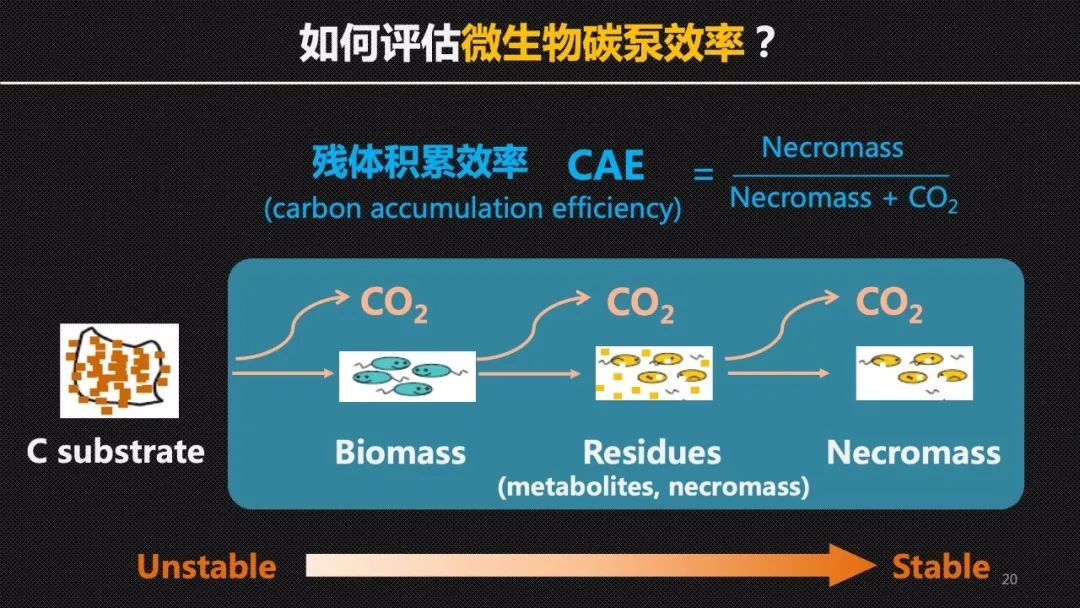
为此我们提出了微生物残体积累效率——CAE的概念,它比较了相同时间内微生物残体积累的总量和呼吸总量的比值。我们可以把微生物的残体看作是合格的加工产品,而那些微生物呼吸产生的二氧化碳就是废品。那么这样一个比值它其实体现了我们微生物或者说加工车间的一个生产效能和产品合格率。
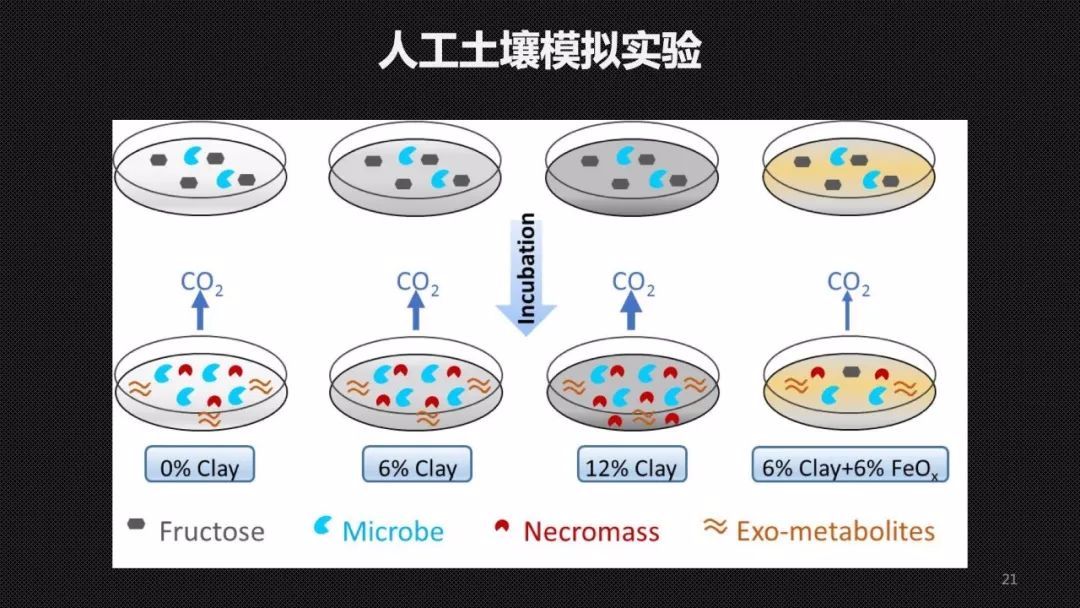
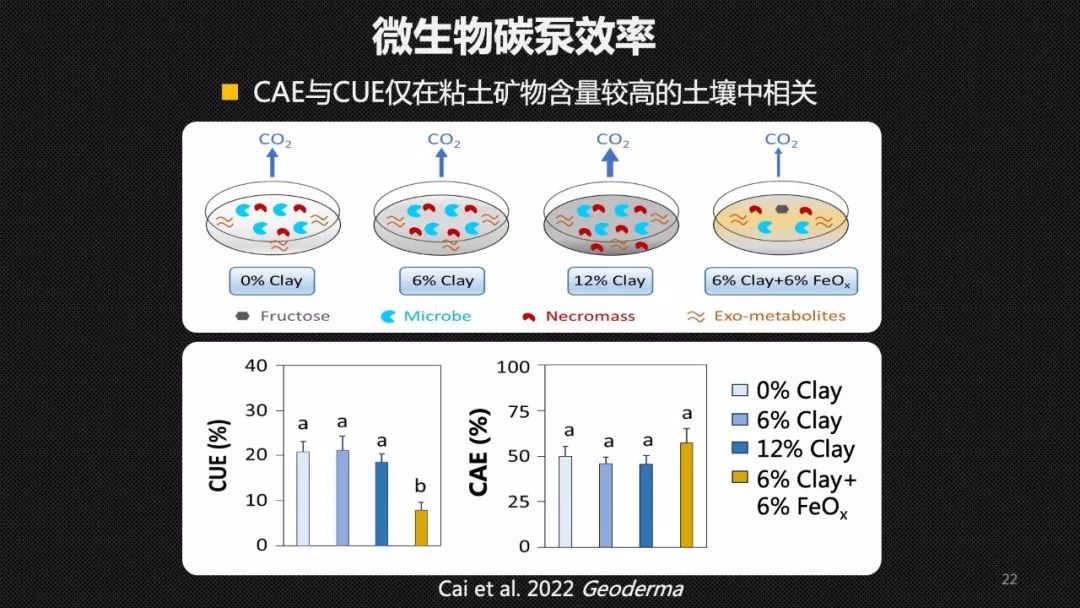
为了验证CAE和CUE之间的关系,我们首先开展了一个人工土壤的模拟实验。我们用添加了不同量的黏土矿物以及铁氧化物的石英砂,作为我们人工土壤的矿物基质。在里面接种了草地微生物的菌液以后, 这个实验进行了4个月,这期间我们定期的去监测土壤微生物的呼吸、产生的生物量、微生物残体指标——氨基糖以及残留果糖的量。因为整个实验中果糖是我们添加的唯一的有机碳的来源,所以我们可以认为在除去了残余果糖以后,土壤中所有的有机碳都经过了微生物的转化,也就是微生物代谢产物的总量。通过这样一个研究,我们发现CAE和CUE只在粘土矿物含量较高,也就是说矿物保护作用比较强的土壤中,它们才正相关,在其他的土壤里面它们可能没有关系。这就证明CAE和CUE确实可能解耦。CAE能够更好地去表征土壤微生物的碳泵效率,去描述这样一个过程。
其实我的实验室里还进行了很多的类似人工土壤的模拟实验,我们去观察土壤碳的一个转化过程,但是由于我们使用的不是真实的土壤,所以并不受青睐。我们的成果不好发表,很多杂志看到人工土壤的研究就直接拒稿,但是我对我备受挫折的研究生说,“这是我非常得意的工作!”
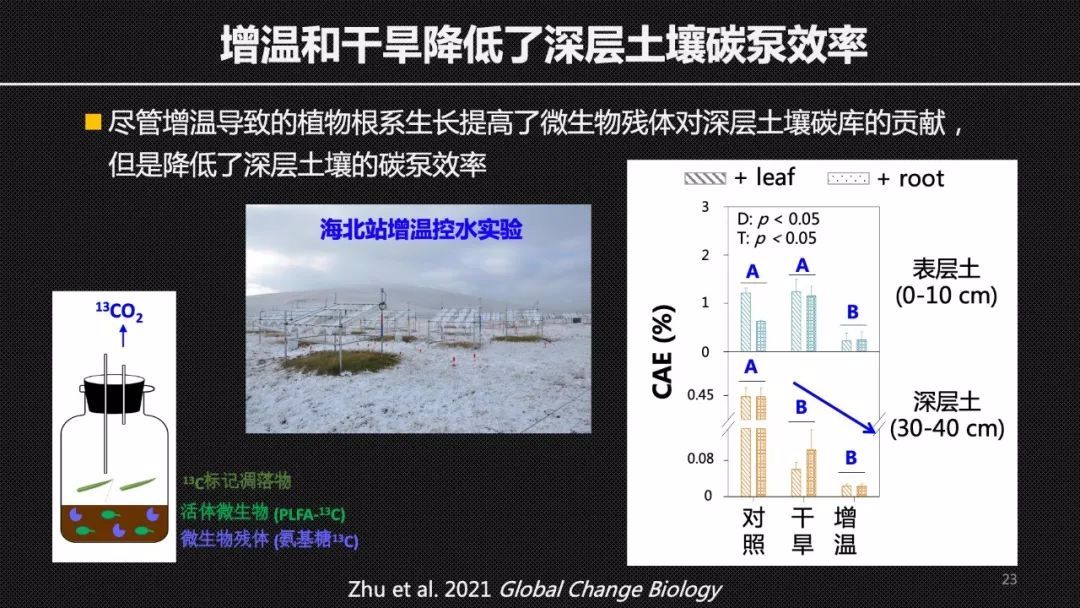
基于我们提出的CAE这样一个参数,我们去评估了海北站的气候变化控制实验下土壤“微生物碳泵”效率对于增温和干旱的响应。我们发现尽管增温增加了深层土壤中植物的根系生长。但是植物根系的生长同时也加剧了植物和微生物对于土壤养分的竞争关系,使得深层土壤中的微生物的氮限制加强,因此降低了深层土壤的土壤“微生物碳泵”效率,也就是CAE。这样一个结果说明,在增温的背景下,由于植物根系的输入提供了更多的原材料,微生物也消耗了更多的植物碳来产生残体,但是同时它的产品合格率下降,产生了更多的废品二氧化碳。这就验证了我们的观点,说明土壤中微生物残体的总量并不能够反映土壤碳封存的一个成效,我们更需要关心的是土壤“微生物碳泵”效率。
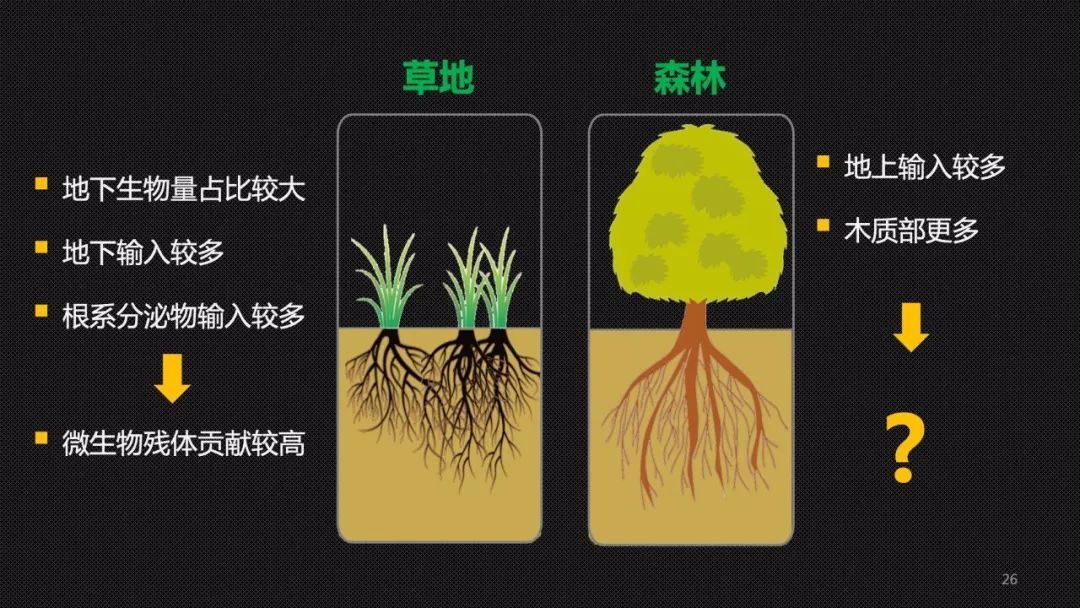
故事讲到这里,大家可能也好奇,是不是所有生态系统的土壤碳积累都由微生物残体来主导?大家也应该猜到答案是否定的。我们刚才的研究聚焦于草地生态系统。草地有什么特点呢?和地上部分相比,草地的地下生物量也就是根系的占比更大,植物将更多的光合作用产物投入到地下部分,其中很大一部分就是根系的分泌物。这样一些溶解态的小分子的化合物很容易被微生物利用消化,进而促进“微生物碳泵”,并且促进残体的积累。换一句话说,比较容易消化和比较好吃的根系分泌物,更加容易提高土壤的“微生物碳泵”效率,提高微生物的加工效能。因此在草地土壤中微生物的残体对于土壤碳库的贡献较高。

相比之下在森林中有更多的地上输入,也就是枯枝落叶,而且枯枝落叶中的木质素的含量较高,对于微生物来说并不好吃。因此,在森林中植物提供的原材料不容易降解,更容易积累。
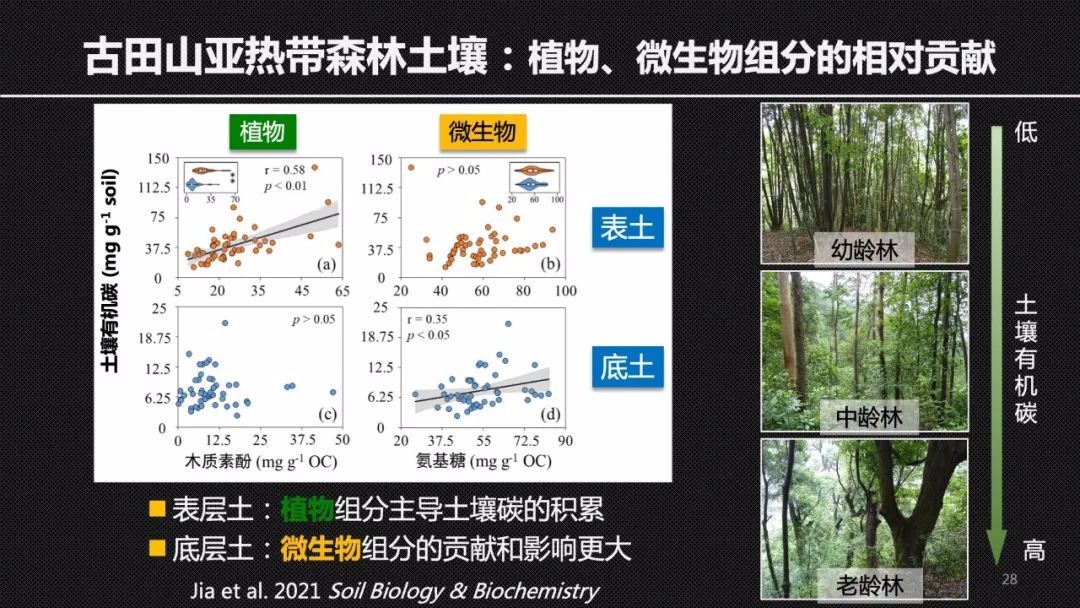
为了验证这样一个想法,我们也对森林土壤进行了研究。这是我的团队在内蒙古的温带草地和古田山亚热带森林采样的照片。大家可以看到和草地相比,森林表层土壤有更多的枯枝落叶覆盖。相应的,古田山的表层土壤有机碳积累的时候,会伴随着木质素,但不是氨基糖的增加,但是在30厘米以下的深层土壤中,土壤有机碳的积累更多的受到微生物残体的影响。

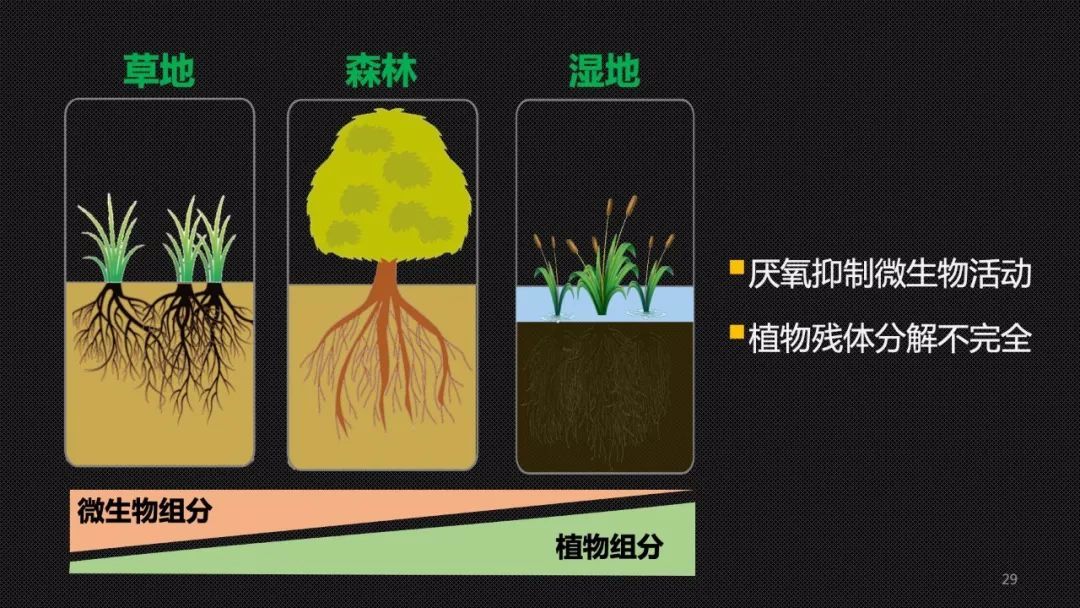
所以在不同的生态系统中,主导土壤碳封存的主力或者关键过程是存在差异的。在微生物活动强烈的草地土壤中,微生物的残体对于草地土壤碳库的贡献比较的大。
因此我们也希望在牧区,大家不要简单地认为围栏封牧、恢复草场就可以有利于碳封存,我们也需要同时关心土壤中微生物的健康状况和它的这样的加工的效能。而在另外一些比如说森林这样的生态系统,甚至更极端的环境,比如说淹水和厌氧的湿地中,微生物的活性受限,那些分解不完全的植物凋落物——枯枝落叶,这个时候就充当了土壤碳封存的主力军。所以我们希望,在适宜森林生长的山区退耕还林这样的一些措施能够继续得到推广。
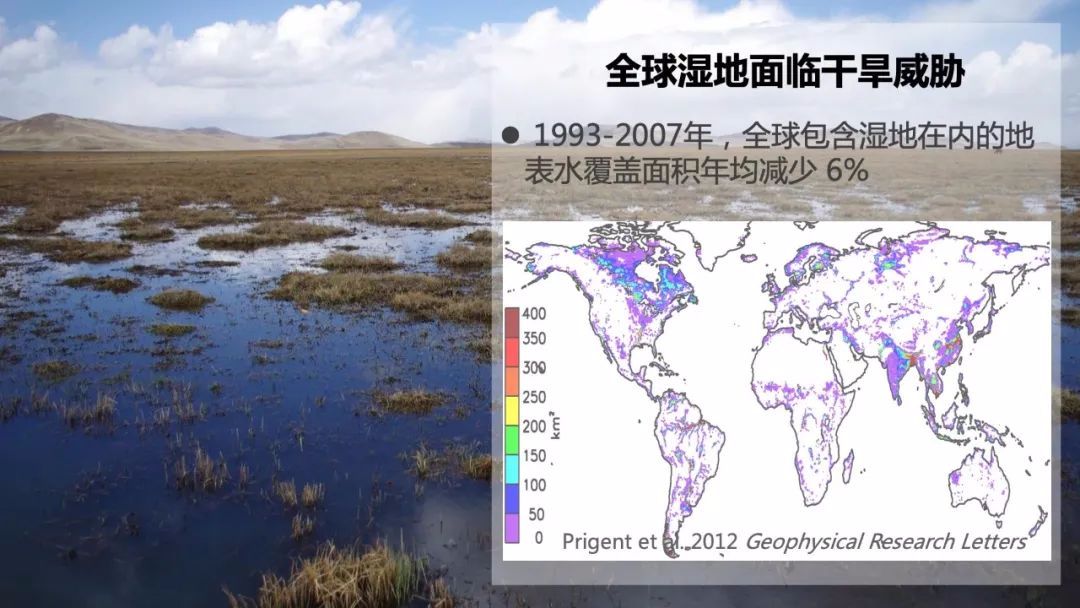
这里我要特别提一下湿地,因为在全球湿地储存了三分之一以上的土壤碳。在地质历史时期上,湿地的退化与扩张对大气二氧化碳浓度的变化有巨大的影响。全球约一半的湿地面临着干旱化的风险。那干旱化会对湿地的土壤碳封存产生怎样的影响呢?
经典的湿地的“酶栓”机制认为,在湿地的土壤有机碳或者说湿地中有机碳的积累,主要得益于厌氧环境对于氧化酶的抑制作用。在湿地排水或者干旱以后,氧气的渗透性会大大提高。这时胞外酶的活性也会大大提高,进而加快有机碳的分解。但是,全球不同湿地对于干旱或者排水的响应并不一致,说明可能还存在其他的调控机制。

在湿地中大量存在的变价金属铁,遇到氧气氧化以后,会形成铁氧化物。这些铁氧化物和酚类、木质素类的这样有机质具有强烈的交互作用,因而可以促进土壤碳的封存。在湿地中铁的转化是否调控了湿地土壤碳库对于干旱的响应?
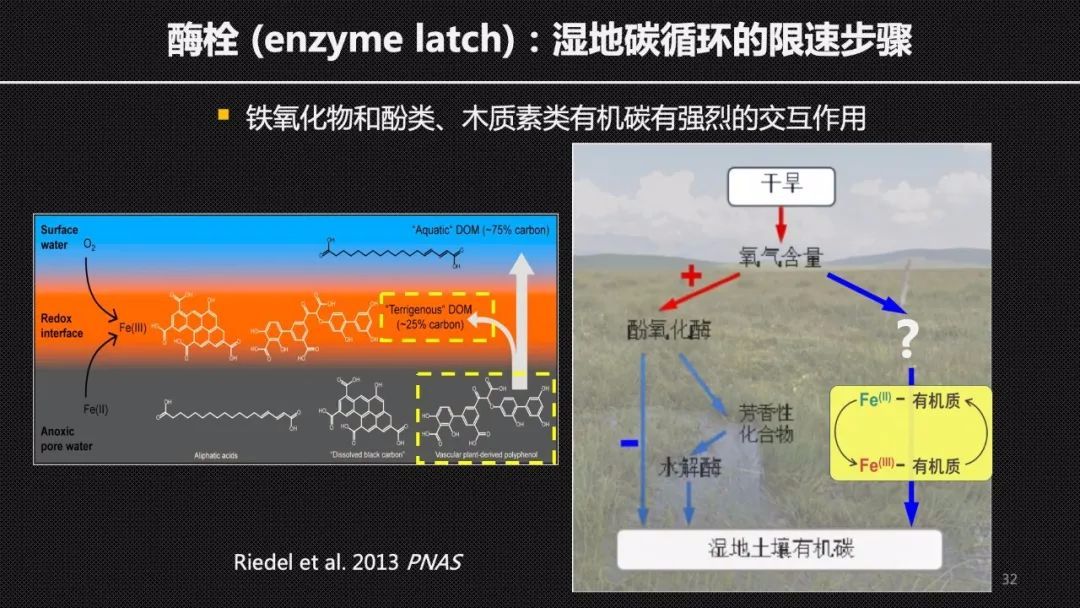
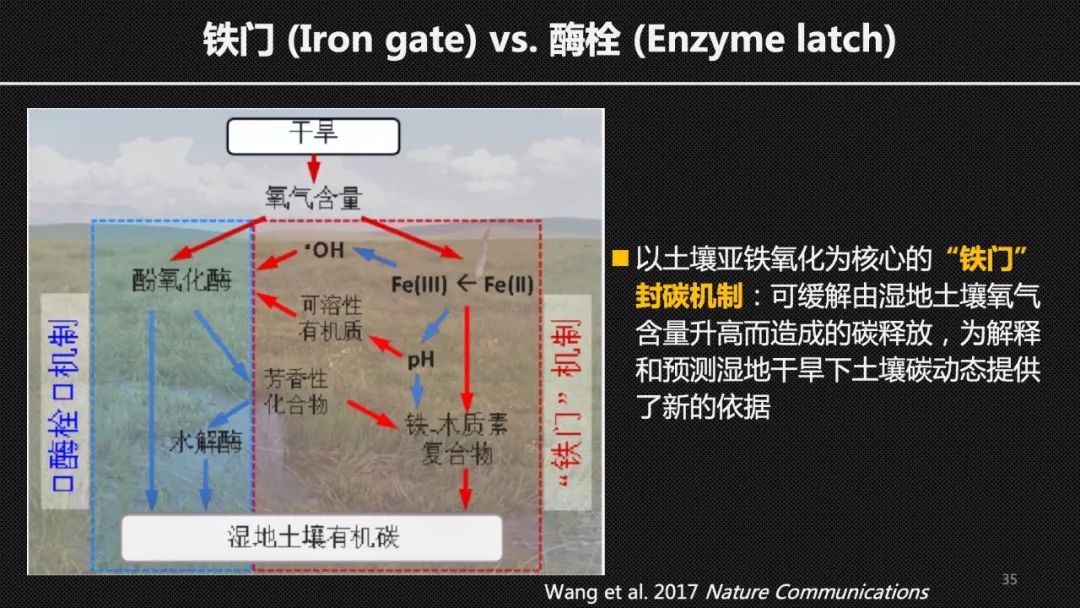
为了验证这样一个假设,我们用控水装置来调控干旱组和对照组的水位,使得干旱组的水位比对照组下降20厘米。在两个生长季的实验以后,我们对土箱中心的土壤进行了采样和分析,我们发现和“酶栓”机制相反。干旱导致的二价铁含量的下降,使得酚氧化酶的活性下降,而不是上升。

为此,我们提出了湿地的“铁门”封碳机制,它可以对抗经典的“酶栓”机制,调控土壤碳,特别是植物碳在干旱下的一个保存,这也表明碳——铁的交互作用可能是湿地中碳封存的一个关键的途径。为我们理解和预测湿地干旱下土壤碳动态提供了新的理论依据。我们的“铁门”机制的论文发表以后也备受关注,而且被国内外的同行积极的验证。
通过以上的研究,我们可以发现土壤碳封存是初级生产者(植物)和转化者(微生物)之间博弈的结果。不同的生态系统也是由独特的气候、地质、动植物、微生物组合而成,它们之间的交互作用和关系的健康维持,对于土壤的生物地球化学循环以及碳封存都具有重要的意义!

当然,目前我们对于土壤封碳的机制以及过程的了解还非常的有限,未来还有许多工作需要开展。科学界也需要开展更多的研究,才能够更好地支持政府决策,通过改善对于森林、草地、湿地、农田的管理,来应对我们日益严峻的降碳的挑战。最后我想说,助力实现“碳中和”的方案就在我们的脚下,土壤封碳尚有潜力,仍需探索!谢谢!

阅读
16174

| 我也说两句 |
| 版权声明: 1.依据《服务条款》,本网页发布的原创作品,版权归发布者(即注册用户)所有;本网页发布的转载作品,由发布者按照互联网精神进行分享,遵守相关法律法规,无商业获利行为,无版权纠纷。 2.本网页是第三方信息存储空间,阿酷公司是网络服务提供者,服务对象为注册用户。该项服务免费,阿酷公司不向注册用户收取任何费用。 名称:阿酷(北京)科技发展有限公司 联系人:李女士,QQ468780427 网络地址:www.arkoo.com 3.本网页参与各方的所有行为,完全遵守《信息网络传播权保护条例》。如有侵权行为,请权利人通知阿酷公司,阿酷公司将根据本条例第二十二条规定删除侵权作品。 |